 科技服務(wù)
科技服務(wù)- 首頁 >> 科技服務(wù) >> 工作動(dòng)態(tài)
工作動(dòng)態(tài)
武漢病毒所/生物安全大科學(xué)中心揭示RNA病毒聚合酶具有一個(gè)與宿主適應(yīng)相關(guān)的特征區(qū)域
RNA病毒編碼的依賴RNA的RNA聚合酶(RNA-dependent RNA polymerase,簡稱RdRP)是一類獨(dú)特的核酸聚合酶,在病毒基因組復(fù)制和轉(zhuǎn)錄過程中發(fā)揮核心作用,是抗病毒藥物研究的熱點(diǎn)靶標(biāo)。病毒的RdRP由于可與其他功能域融合或與其他病毒蛋白共折疊,其整體結(jié)構(gòu)多樣性較高,但其催化核心區(qū)的三維結(jié)構(gòu)則較為保守,因此RdRP兼具多樣性和保守性。由于RNA病毒的宿主范圍幾乎囊括了所有細(xì)胞形式的生命體,在與不同宿主的共同進(jìn)化過程中,RdRP作為RNA病毒最保守的蛋白,其與病毒的宿主適應(yīng)性之間的關(guān)聯(lián)并不清晰。
黃病毒包括乙型腦炎病毒(Japanese encephalitis virus,簡稱JEV)、登革病毒(dengue virus,簡稱DENV)、寨卡病毒、蜱傳腦炎病毒(tick-borne encephalitis virus,簡稱TBEV)等多種人類致病病原,是一類分布廣泛、種類繁多的單股正鏈RNA病毒,與丙型肝炎病毒和經(jīng)典豬瘟病毒同屬于黃病毒科。黃病毒大多由吸血節(jié)肢動(dòng)物如蚊和蜱作為媒介傳播,可引起人類腦炎或出血性疾病,對人類健康構(gòu)成重大威脅。黃病毒的RdRP位于病毒編碼的非結(jié)構(gòu)蛋白NS5的羧基端,與氨基端的甲基轉(zhuǎn)移酶(MTase)形成天然融合體。此前蚊傳黃病毒的NS5已有多個(gè)三維結(jié)構(gòu)報(bào)道,包括全長蛋白、MTase區(qū)和RdRP區(qū)的結(jié)構(gòu),而蜱傳黃病毒NS5蛋白的全長結(jié)構(gòu)及RdRP三維結(jié)構(gòu)則未獲解析。
中國科學(xué)院武漢病毒研究所龔鵬研究員團(tuán)隊(duì)長期從事病毒RdRP的催化與調(diào)控機(jī)制研究,該團(tuán)隊(duì)此前分別解析了JEV和DENV的NS5全長蛋白晶體結(jié)構(gòu)(Lu and Gong, PLoS Pathog 2013; Wu et al., PLoS Pathog 2020),并與武漢病毒所張波研究員團(tuán)隊(duì)合作,系統(tǒng)性揭示了NS5的構(gòu)象多樣性和保守性以及MTase調(diào)控RdRP的分子機(jī)制(Li et al. PLoS Negl Trop Dis 2014; Wu, et al. J Virol 2015; Wu et al., PLoS Pathog 2020)。該團(tuán)隊(duì)近期解析了分辨率為1.9 埃的TBEV MTase晶體結(jié)構(gòu)(PDB號(hào)7D6M,圖1A)和分辨率為3.2埃的TBEV RdRP的晶體結(jié)構(gòu)(PDB號(hào)7D6N,圖1B),獲得了首個(gè)蜱傳黃病毒RdRP的三維結(jié)構(gòu)信息。通過黃病毒RdRP的序列分析以及與結(jié)構(gòu)已知的蚊傳黃病毒RdRP的結(jié)構(gòu)進(jìn)行比較,發(fā)現(xiàn)在RdRP保守的催化基序(motif )B和C之間存在一個(gè)值得關(guān)注的區(qū)域(以下稱B-C連接區(qū))。該區(qū)域位于RdRP手掌區(qū)的底部且暴露于蛋白表面,且在黃病毒屬的不同宿主分類中具有明顯的宿主相關(guān)多樣性(圖1B及圖2)。
該團(tuán)隊(duì)設(shè)計(jì)了TBEV和JEV病毒間B-C連接區(qū)的替換突變,在酶學(xué)水平證實(shí)突變對RdRP催化功能不構(gòu)成本質(zhì)影響(圖3,A-B)。團(tuán)隊(duì)通過與武漢病毒所王漢中研究員/鄭振華研究員團(tuán)隊(duì)和張波研究員團(tuán)隊(duì)合作,在細(xì)胞水平分別評(píng)測了突變對TBEV和JEV的影響,結(jié)果表明在兩種病毒體系中突變后的病毒不能維持在細(xì)胞中的增殖(圖3,C-D)。這些結(jié)果提示B-C連接區(qū)很可能參與了RdRP催化以外與病毒增殖相關(guān)的重要過程。通過對正鏈、負(fù)鏈和雙鏈RNA病毒中的代表性RdRP的B-C連接區(qū)進(jìn)行結(jié)構(gòu)與序列的系統(tǒng)分析,在RNA病毒大家族中進(jìn)一步發(fā)現(xiàn)該區(qū)域在結(jié)構(gòu)和序列長度方面具有較高的多樣性(圖4),提示RdRP的B-C連接區(qū)很可能是RNA病毒共有的一個(gè)宿主適應(yīng)熱點(diǎn)區(qū)域,此項(xiàng)研究為病毒RdRP的調(diào)控機(jī)制研究及RdRP相關(guān)的宿主適應(yīng)研究提供了重要線索。
此項(xiàng)研究主要受到國家重點(diǎn)研發(fā)計(jì)劃項(xiàng)目“畜禽重要病原共感染與協(xié)同致病機(jī)制研究”(2018YFD0500100,項(xiàng)目負(fù)責(zé)人為中國農(nóng)業(yè)科學(xué)院上海獸醫(yī)研究所丁鏟研究員)和NSFC面上項(xiàng)目(31670154; 32070185)的支持。博士研究生楊婕妤和博士后景旭平為論文的共同第一作者,主要完成了結(jié)構(gòu)與酶學(xué)研究工作,龔鵬、鄭振華和張波為共同通訊作者,王漢中/鄭振華團(tuán)隊(duì)的實(shí)驗(yàn)師易文富、碩士研究生姚琛和張波團(tuán)隊(duì)的博士后李曉丹分別完成了TBEV和JEV病毒學(xué)研究工作,相關(guān)論文近期于Nucleic Acids Research(《核酸研究》)上在線發(fā)表,原文鏈接為: https://academic.oup.com/nar/advance-article/doi/10.1093/nar/gkaa1250/6066638?guestAccessKey=c6d13ee0-287e-4d7b-97dc-01d9456423e7
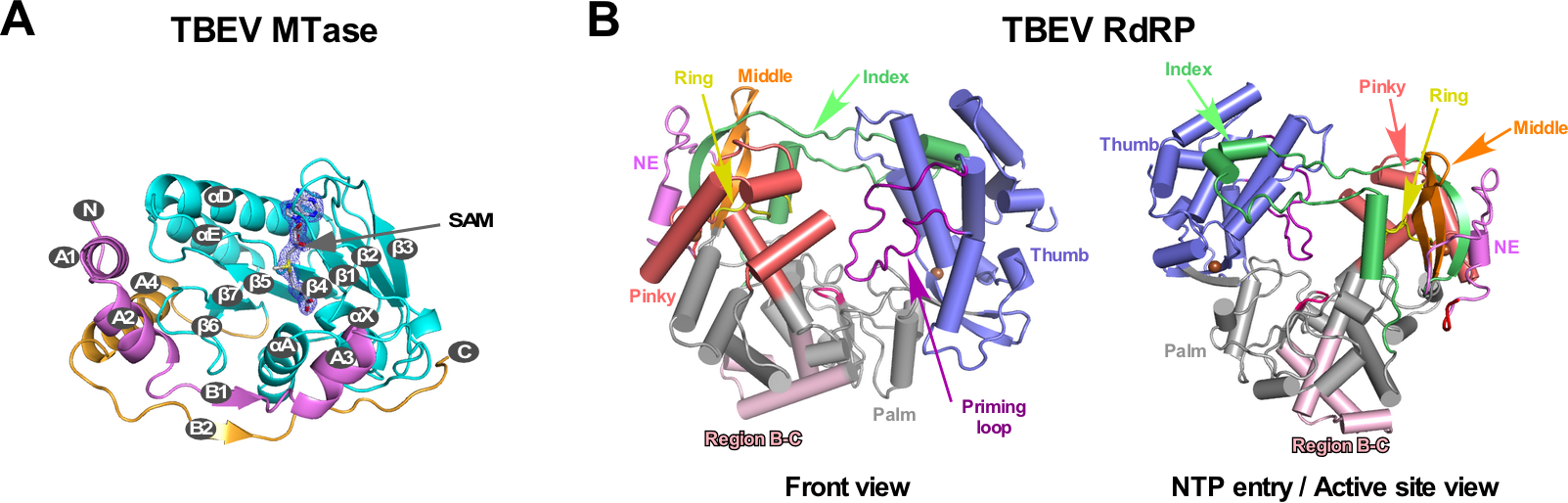
圖1. TBEV MTase和RdRP的晶體結(jié)構(gòu)。A)MTase結(jié)構(gòu)中結(jié)合了一個(gè)甲基供體SAM分子。B)RdRP由拇指、手指和手掌三個(gè)結(jié)構(gòu)域組成,B-C連接區(qū)(淺粉色)位于手掌區(qū)底部。
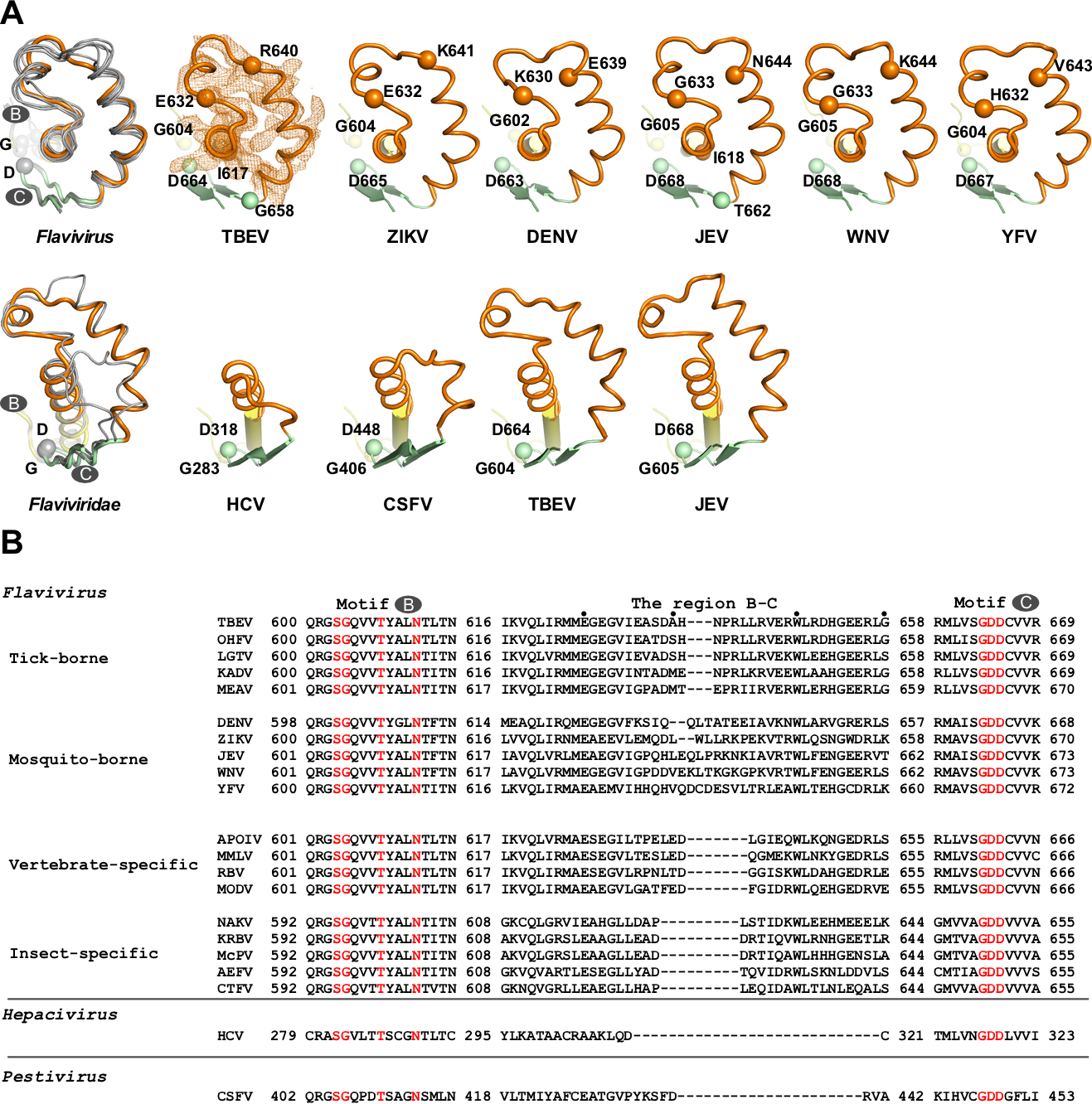
圖2. 黃病毒科RdRPs中B-C連接區(qū)結(jié)構(gòu)和序列的多樣性。A)第一排中TBEV屬于蜱傳黃病毒,其余屬于蚊傳黃病毒。第二排中TBEV和JEV屬于黃病毒屬,而HCV和CSFV分別屬于黃病毒科中的肝炎病毒屬和瘟病毒屬。其中motif B、B-C連接區(qū)和motif C分別用黃色、橙色和淺綠色表示。B)黃病毒NS5中B-C區(qū)域的序列比對。根據(jù)宿主的不同,黃病毒可以分為蚊傳、蜱傳、僅在昆蟲中傳播和僅在脊椎動(dòng)物中傳播四類。

圖3. 病毒種間B-C連接區(qū)替換突變影響病毒增殖。A-B)將連接區(qū)替換為JEV序列的TBEV NS5表現(xiàn)出與野生型聚合酶相似的依賴引物(A)和從頭合成(B)活性。C-D)在細(xì)胞水平上TBEV和JEV的連接區(qū)替換突變體均不能實(shí)現(xiàn)病毒增殖。
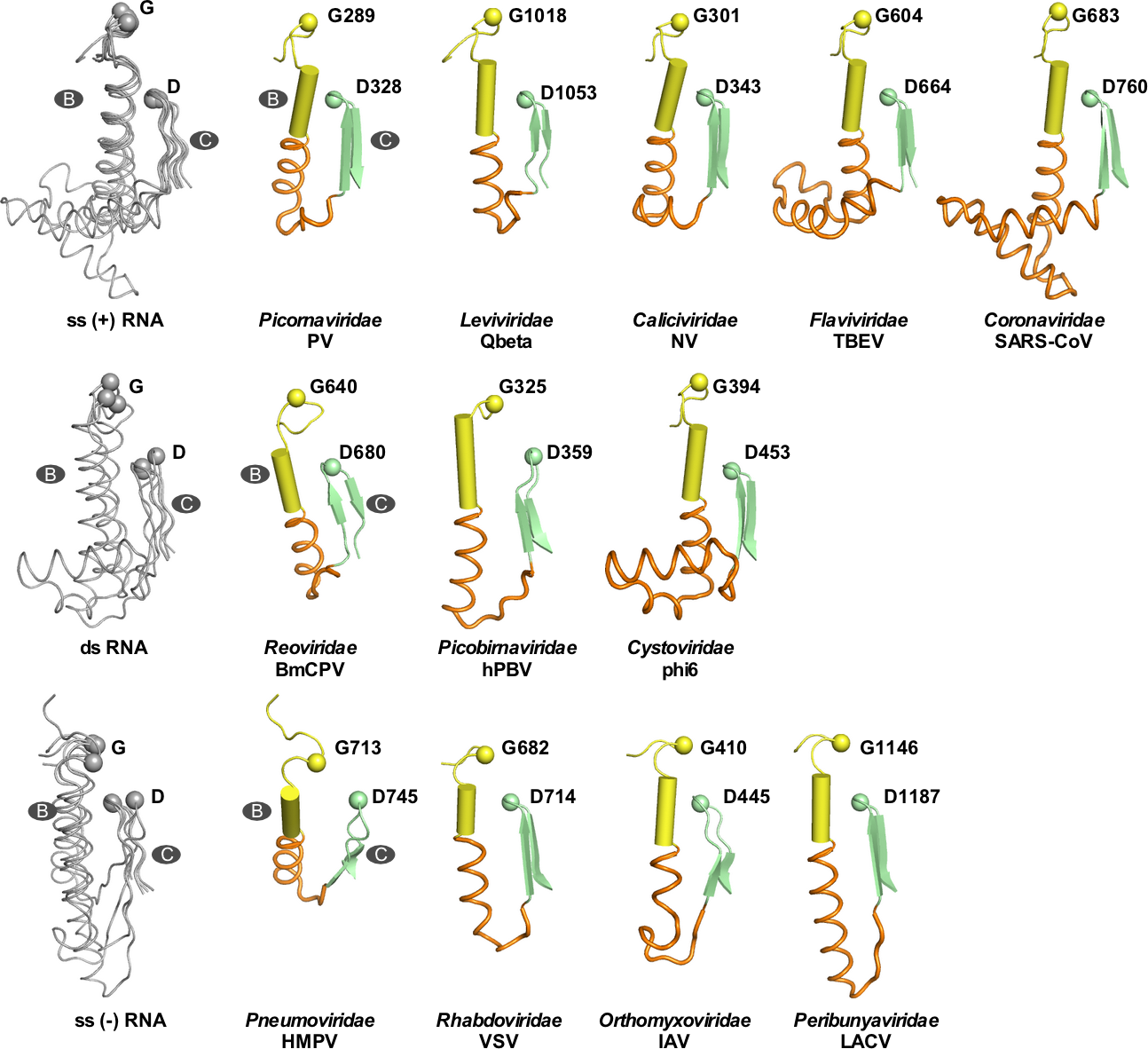
圖4. 病毒RdRP的B-C連接區(qū)展現(xiàn)出較高的多樣性。上中下三行分別為正鏈、負(fù)鏈和雙鏈RNA病毒代表性RdRP的連接區(qū)結(jié)構(gòu)。每個(gè)彩色結(jié)構(gòu)分別代表一個(gè)RNA病毒科。其中B-C連接區(qū)用橙色顯示。左側(cè)為多結(jié)構(gòu)疊印圖(灰色)。